Compatibilidad en la polinización entre variedades de manzana para sidra
La elaboración de sidra de manzana tiene una larga tradición en Gipuzkoa que se remonta al menos hasta la Edad Media (Fundación Hazi, 2016). Gracias a ello, en la actualidad todavía se cultivan más de cien variedades tradicionales, si bien la mayoría de la producción se concentra en 24, que son además recomendadas por la D.O. Euskal Sagardoa/Sidra natural del País Vasco (cita), que ampara desde 2016 la producción de sidra de calidad a partir únicamente de manzanas de variedades autóctonas.
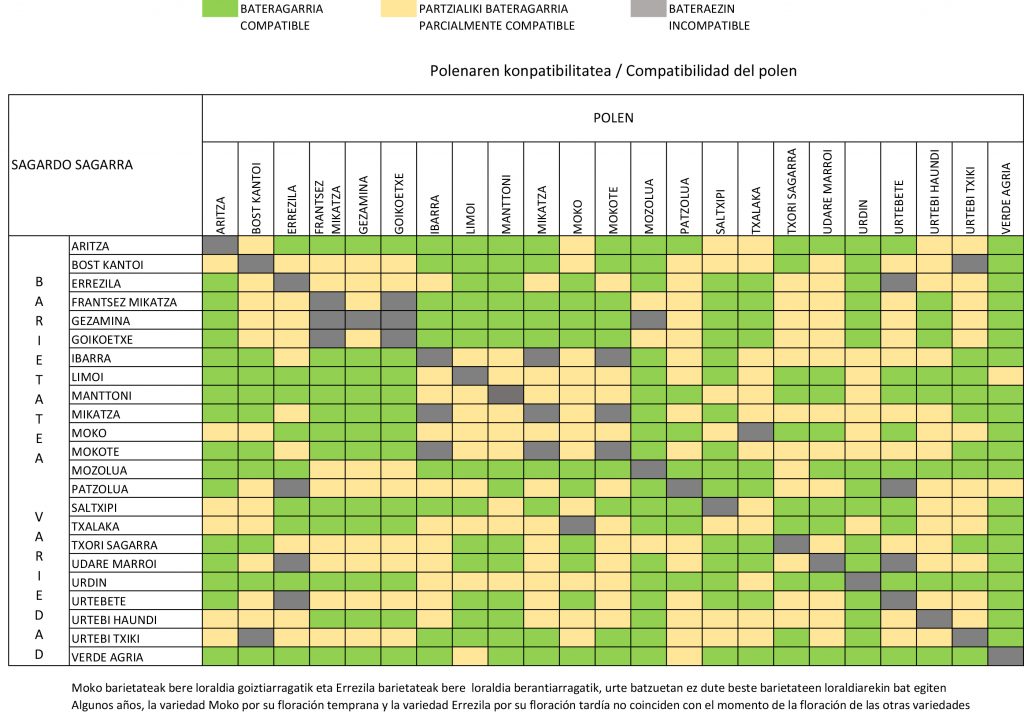
Dependiendo de la combinación de S-alelos entre la variedad polinizante y la polinizada, la compatibilidad puede ser total, parcial, o nula
A pesar de la alta y reconocida calidad de la producción, la rentabilidad de los manzanales de sidra guipuzcoanos se ve en muchas ocasiones comprometida por una escasa productividad (cita sagarlan/diputación). Son varias las razones por las que puede explicarse, entre las que se encuentran la propia naturaleza fuertemente vecera de muchas de estas variedades, la climatología desfavorable durante las épocas de floración-cuajado, un manejo agronómico inadecuado y, especialmente, que se desconoce muchos aspectos de la biología reproductiva de las variedades. Más allá de la época de floración, se desconoce su grado de intercompatibilidad genética (S-alelos) y la calidad y viabilidad del polen, lo que dificulta la correcta elección de polinizadores en el diseño de las plantaciones.
El manzano presenta un sistema de auto-incompatibilidad determinado genéticamente por una región del genoma denominada locus S, cuya función evolutiva es la de impedir los efectos negativos de la endogamia (Broothaerts, 2003; Sheick et al., 2018). Debido a este sistema, si el S-alelo del grano de polen coincide con alguno de los S-alelos del estilo de la flor receptora, el grano será incapaz de crecer y orientarse en el estilo hasta llegar al óvulo. Dependiendo de la combinación de S-alelos entre la variedad polinizante y la polinizada, la compatibilidad puede ser total, parcial, o nula (incompatibles). La compatibilidad total se da cuando la variedad polinizante tiene todos sus S-alelos diferentes a la polinizada, la parcial cuando sólo uno lo es diferente, y en el caso de que todos los S-alelos coincidan se da la incompatibilidad. Hasta el momento, se han identificado más de 50 S-alelos en manzano, tanto en variedades cultivadas como silvestres. Esto supone una alta diversidad, aunque en la práctica tan sólo diez (S1, S2, S3, S5, S7, S9, S10, S20, S24 y S28) suponen más del 80% de los S-alelos presentes en las más de 300 variedades analizadas en la literatura (Broothaerts, 2003; Dreesen et al., 2010; De Franceschi et al., 2018; Halász et al., 2011; Sheick et al., 2020). Esto se debe a que los estudios se han realizado en su gran mayoría para variedades modernas y comercialmente importantes de mesa, en las que unas pocas (como Golden, Gala, Jonathan, Fuji o Red Delicious) son parentales muy populares en los programas de mejora genética.
Por otra parte, la calidad y viabilidad del polen varía entre especies y variedades de manzano. Es ampliamente conocida la alta calidad de Granny Smith como polinizador, así como la de variedades silvestres de Malus floribunda, (como Evereste, Golden Gem o Baugene) y que las variedades triploides como Reineta o Jonagold suelen ser malos polinizadores a causa de la escasa viabilidad de su polen.
En este trabajo se presentan los grupos de auto(in)compatibilidad genética (S-alelos) y la viabilidad del polen de las principales variedades tradicionales de sidra cultivadas en Gipuzkoa.
Material y métodos
El estudio se ha realizado los años 2020 y 2021 sobre las 25 variedades de manzana de sidra tradicional recomendadas por la D.O. Euskal Sagardoa/Sidra natural del País Vasco, entre las que se encuentran 19 variedades diploides y 5 triploides. Los árboles de los que se tomaron las muestras estaban plantados en la finca Otalarrea, perteneciente a la Diputación Foral de Gipuzkoa o en el Banco de Germoplasma de la UPNA. Se incluyeron también ocho variedades modernas, con S-alelos conocidos, como referencia para garantizar el buen resultado de los análisis.
La caracterización genética de los S-alelos se realizó mediante un único par de cebadores inespecíficos (De Franceschi et al., 2016, 2018; Sheick et al., 2018), con un proceso que permite detectar todas las variantes de S-alelos presentes en la muestra y comparar su secuencia con una base de datos internacional (NCBI, Nucleotide Blast). Los S-alelos para los que se encuentra una equivalencia en la base de datos quedan identificados, y si no se encuentra equivalencia, se trata de un S-alelo nuevo, no descrito hasta el momento.
Dado que se ha trabajado con material tradicional, era previsible encontrar nuevos S-alelos, ya que los trabajos realizados hasta la fecha se han centrado mayoritariamente en variedades de mesa relevantes comercialmente o en mejora. En los casos en que se encontraron equivalencias, se realizó una confirmación adicional empleando marcadores específicos.
En total, se detectaron 11 S-alelos ya conocidos en las variedades estudiadas, que fueron identificados como S1, S2, S3, S5, S7, S11, S20, S23, S24, S26 and S28. Se detectó un alelo adicional en la variedad Verde Agria para el que no se encontró equivalencia en la base de datos internacional, y que podría corresponder a un nuevo S-alelo no descrito previamente. Dos S-alelos, presentes en las variedades triploides Urtebi Haundi y Mozolua codificaron como alelos nulos y no pudieron ser identificados. Como ya se ha indicado, el hallazgo de nuevos S-alelos no era inesperado, dado que hasta la fecha la mayor parte de los estudios se han centrado en variedades modernas (Broothaerts et al., 2004; Dreesen et al., 2010; De Franceschi et al., 2018), y cuando se han realizado sobre material tradicional o local ha sido común encontrar alelos que hasta ese momento no se había descrito (Broothaerts, 2003; Dreesen et al., 2010). La frecuencia con la que los S-alelos han aparecido en las variedades tradicionales analizadas ha sido muy diferente de la encontrada en estudios previos (Broothaerts et al., 2004; Dreesen et al., 2010; De Franceschi et al., 2018; Halász et al., 2011) hechos sobre un gran número de variedades, tanto modernas como tradicionales. S3, S5, S26 and S28 fueron comunes (>10% de los S-alelos) o muy comunes (>15%) en nuestra muestra. Es de destacar la alta frecuencia con la que se detectó al alelo S26, ya que apareció en nueve de las variedades. A este S-alelo se le considera extremadamente infrecuente en manzano cultivado (M. × domestica), pero aparece con frecuencia en las variedades silvestres como M. sylvestris and M. floribunda (Broothaerts, 2003; Dreesen et al., 2010). De todas las selecciones comercializadas, en este momento solo se conoce el genotipo de Malus Evereste®, que es S20 S26. Los dos alelos que porta son extremadamente raros en manzano de mesa, pero no en el manzano tradicional estudiado: en las variedades de este estudio se ha encontrado una variedad (Haritza) con el S20, y nueve (Limoi, Moko, Txalaka, Reineta Encarnada, Manttoni, Urdin, Mikatza, Mokote e Ibarra) que portan el S26. Esto hace que Evereste sea una variedad parcialmente compatible con todas ellas, ya que solo la mitad del polen que produce es capaz de fecundarlas. Si coinciden en época de floración, Evereste sería un polinizador adecuado, pero no excelente como lo es para las variedades de mesa modernas. La situación podría ser similar para los otros manzanos silvestres comercializados como polinizadores, pero en ellos se desconoce su genotipo, y hacerlo sería un requisito indispensable a la hora de considerarlos o recomendarlos como potenciales polinizadores de las variedades de sidra en la D.O. En conjunto, estos resultados serían un indicador adicional de la existencia de una “población ibérica” diferenciada del resto del manzano tradicional europeo, que ya había sido detectado mediante análisis de estructura genética (Pereira-Lorenzo et al., 2017).
Las variedades quedaron clasificadas en 18 grupos de incompatibilidad (Tabla 1), de los que 13 corresponden a variedades diploides. Cada variedad triploide tuvo su propio S-genotipo, pero en el caso de las diploides, en cinco casos hubo dos o más variedades que compartían genotipo y, por tanto, eran completamente incompatibles entre sí.
Conclusiones
Se han identificado once S-alelos y 18 grupos de incompatibilidad entre las 24 variedades tradicionales de sidra cultivadas en Gipuzkoa y recomendadas por la D.O. Uno de los S-alelos no había sido descrito con anterioridad, y las frecuencias con que los S-alelos
aparecieron fueron muy distintas a lo que se observa en otras variedades locales europeas o en las variedades modernas. Estos resultados constituyen una prueba más de la existencia de una “población ibérica” de manzano autóctono, que ya había sido identificado mediante análisis de estructura genética. La viabilidad del polen fue generalmente buena en las variedades diploides y mala en las triploides. Sin embargo, Urtebi Haundi (triploide) ha mostrado una viabilidad bastante buena y Urtebete y Errezila (diploides) muy baja. Los resultados obtenidos en el estudio permitirán mejorar el diseño de nuevas plantaciones de manzano de sidra tradicional en la región, introducir mejoras en las ya implantadas, así como mejorar la rentabilidad de los manzanales de sidra plantados con estas variedades.
Autor: Carlos Miranda (UPNA)
Fuente: Anuario Sagardoaren Lurraldea 2024
La entrada Compatibilidad en la polinización entre variedades de manzana para sidra aparece primero en Sagardoaren Lurraldea.
Share on Facebook Share on Twitter Share on Pinterest